细胞衰老概念
- 细胞衰老现象最早在1961年被报道。
- 1961 年,美国生物学家Leonard Hayflick在体外培养正常人的成纤维细胞时发现,即使给予细胞生长最适宜的条件,细胞分裂至一定代数时也会发生衰竭, 也就是细胞周期进入了一种“不可逆”的停滞状态。基于此种现象,Hayflick首次提出了细胞衰老的概念(cellular senescence)。而这种细胞的增殖能力的极限被称为Hayflick极限(Hayflick limit)。
- 机制研究表明,这种现象是因为细胞随着复制次数越来越多,从而导致端粒的缩短和端粒酶活性的降低。随着研究的深入,研究人员发现,有很多因素都可能导致细胞衰老,除了端粒缩短而导致的细胞衰老之外,其他的体外刺激也同样可以诱导细胞发生衰老现象。
- 常见的体外刺激包括活性氧自由基的产生、癌基因的表达或抑癌基因的失活、化疗和放疗以及诱导细胞发生重编程等刺激。
细胞衰老分类
- 细胞衰老(cellular senescence)的定义:随着时间的推移或面临外界应激压力时,细胞的正常生理功能和增殖能力发生逐渐地衰退,从而脱离细胞周期的过程。整个细胞衰老过程和机体的衰老及疾病等,都具有重要的关联
按照细胞衰老的诱发因素进行分类
- 端粒诱导的细胞衰老(Telomere-induced senescence)
- 癌基因诱导的衰老(Oncogene-induced senescence,OIS)
- 氧化应激诱导的细胞衰老(Oxidative stess-induced senescence)
- 治疗诱导的细胞衰老(Treatment-induced senescence,TIS)
- 重编程诱导的细胞衰老(Reprogramming-induced senescence,RIS)
- …
按照细胞衰老的发生机制进行分类
生理性原因引起的细胞衰老/复制性细胞衰老Replicatice Senescence
- 正常培养的细胞之所以发生细胞衰老,是因为细胞多次增殖分裂之后,端粒缩短,细胞达到了正常细胞分裂的极限,也就是Hayflick 极限
过早衰老Premature Senescence/加速衰老Accelerated Senescence
- 其他诱因导致的细胞衰老,常常是由于各种各样的其他刺激(例如,癌基因突变,氧化应激刺激,治疗刺激等等),从而导致细胞在达到 Hayflick极限之前,就开始了的,细胞的衰老过程。
常见的细胞衰老现象
复制性衰老
- 复制性衰老现象是由于细胞因为不断地分裂导致端粒缩短,从而诱导细胞发生衰老的现象。
- 端粒是位于染色体末端的DNA重复序列(在脊椎动物细胞中,其序列是 TTAGGG)以及结合在这段DNA重复序列之上的核蛋白复合物所共同组成的复杂结构。
- 端粒的作用是在染色体末端维持基因组的稳定性。正常细胞端粒的长度会随着DNA 的复制而不断地缩短。当端粒DNA缩短到一定程度时,细胞会自动开启衰老程序,这个过程就被称之为端粒缩短诱导的细胞衰老Telomere-induced senescence,也可以被称之为复制性细胞衰老Replicative senescence。
- 端粒缩短和功能异常可能会引起细胞DNA损伤应答(DNA damageresponse, DDR) , 导致细胞衰老的发生;相反,如果过表达端粒逆转录酶Telomere reversetranscriptase/TERT则可以推迟或者避免正常细胞发生复制性衰老,从而使得细胞产生永生化。
过早衰老
- 端粒酶活性的增加,并不能阻断癌基因诱导的细胞衰老或者其他应激反应引起的细胞衰老。这说明,由端粒诱导的复制性细胞衰老现象和由应激反应而诱导的过早衰老现象,其下游的机制是不同的。
- 常见的过早衰老现象包括癌基因诱导的细胞衰老、氧化应激诱导的细胞衰老、治疗诱导的细胞衰老和细胞重编程诱导的细胞衰老现象。
癌基因诱导的细胞衰老
- 这种现象虽然被称为癌基因诱导的细胞衰老现象,但是抑癌基因的失活诱导的细胞衰老现象,也被包含在这个概念之中,请不要被名字所迷惑。
- 1997年,有学者将癌基因 Ras 分别转入人和小鼠的成纤维细胞,结果这些转入了癌基因的细胞大部分都变得扁平,并且发生了永久性的G1期细胞周期阻滞现象。除此之外,细胞内还发生了p53、p16等蛋白过度积累的情况。这种现象就是后来被命名为癌基因诱导的细胞衰老现象(Oncogen-induced senescence,OIS),在细胞内过表达Ras 也是诱导OIS最常用的细胞模型。
- 此外,其他的癌基因,例如c-Myc、BRAF^E600 等癌基因的活化,也被报道在特定条件下可以诱导细胞发生衰老。除了癌基因的激活之外,抑癌基因PTEN的失活,也可以诱导细胞发生衰老。
- 有观点认为癌基因诱导的细胞衰老过程中,同样激活了DNA损伤反应(DNA damage response,DDR),从而导致了细胞衰老现象的发生。另有观点认为,癌基因激活后(或者癌基因失活后),还可以在某些处于癌前病变的细胞内,诱导端粒功能的紊乱,加速了细胞衰老的产生。
- 随着研究不断地深化,癌基因诱导的细胞衰老现象,已经被认为是体内抵抗肿瘤发生的重要屏障,细胞衰老和肿瘤的发生/发展现象之间具有复杂的相互关联。
氧化应激诱导的细胞衰老
- 在 1997 年,有研究人员在培养的成纤维细胞内加入了H2O2、后观察到细胞受到H2O2刺激,出现氧化应激反应之后,出现类似衰老的现象。在研究由癌基因Ras过表达而诱导的细胞衰老过程中,研究人员也发现在不同氧气浓度条件下,Ras诱导细胞发生衰老现象的能力也是不同的。以上两种现象促使研究人员意识到氧化应激反应对细胞衰老现象具有促进作用。
- 产生氧化应激的主要原因是细胞内游离的活性氧自由基ROS。在细胞培养基中加入H2O2,或者通过抑制超氧化物歧化酶SOD1,从而间接在细胞内提高ROS的浓度,都可以诱导细胞衰老的发生。
- 活性氧自由基ROS 可以通过两种途径引起细胞衰老
- 第一种方式是ROS氧化细胞内的DNA、脂类物质、蛋白等生物大分子,从而破坏细胞的结构和功能,诱导细胞发生衰老
- 第二种方式是ROS 直接作为细胞内的第二信使,调节细胞内的信号传导通路,诱导细胞发生衰老现象。
治疗诱导的细胞衰老
- 研究发现,肿瘤的放化疗都可以诱导细胞发生衰老现象,常见的放化疗处理有顺铂处理、阿霉素处理或者伽马射线等等。
- 有意思的是,当化疗采用较低的药物浓度或者放疗采用较低的放射强度时,这样的放化疗处理能诱导细胞发生衰老现象;反之,如果采用较高的化疗药物浓度或者较强的放射强度往往诱导细胞发生凋亡。
- 放化疗治疗诱导的细胞衰老的主要原因也是导致了细胞内DNA 的损伤,而部分化疗药物引起细胞内ROS的产生或者因放化疗而在细胞内导致了端粒酶活性受到抑制,从而同样参与诱导细胞衰老,并且有可能加速了细胞的衰老过程。
- 有学者特别提出,在机体接受放化疗治疗之后,虽然肿瘤细胞发生衰老,退出细胞周期,丧失了增殖能力,但是这些细胞具有潜在的致瘤性,而且肿瘤细胞在发生衰老现象后,可能会通过SASP现象(衰老相关分泌表型senescence associated secretion phenotype)促进肿瘤的进展或者转移。
细胞重编程诱导的细胞衰老
- 在 2006 年,日本京都大学的山中伸弥教授带领的科研团队从胚胎干细胞相关的很多候选基因中筛选出四个基因:Oct4、Sox2、KLF4 和 c-Myc,然后通过逆转录病毒将这四个基因导入小鼠胚胎成纤维细胞之内,结果发现这四个基因导入后的胚胎成纤维细胞会发生重编程现象,形成类似胚胎干细胞的细胞,被称为诱导多能干细胞(induced pluripotent stem cell, iPSC)。
- iPSC 的产生效率非常低下,原因是细胞衰老现象是抑制重编程效率的重要原因之一。如果抑制细胞衰老相关通路,可以显著地提高 iPSC 的重编程效率。
- 还有研究人员发现,在产生iPSC的过程中导入了这四个重编程因子之后会诱导细胞发生衰老现象,从而阻碍iPSC 的产生效率,这种现象就被称为重编程诱导的细胞衰老。
细胞衰老的基本特征
细胞周期阻滞
- 细胞衰老最基本的特征之一,就是细胞长久地处于细胞周期的阻滞现象或者永久退出细胞周期,因此细胞周期阻滞现象是在体外鉴定细胞衰老最基本也是最不可或缺的指标之一。
- 检测细胞是否发生细胞周期阻滞的方法有很多。例如,通过检测 Ki67 反应细胞的增殖状态;通过Brdu 渗入实验检测细胞内DNA的复制合成情况,或者通过流式细胞术结合 PI 染色的方法检测细胞在不同细胞周期内的分布情况等。这些都是常用的用于检测细胞周期阻滞的方法,在我们之前的内容中,都有所涉及。
细胞形态的改变
- 细胞在衰老后的形态学变化只能作为判断细胞衰老的辅助性证据
- 发生衰老的细胞和正常细胞相比,在形态上具有显著不同的特征。例如,衰老细胞通常会表现出细胞变得扁平,整个细胞的胞体和细胞核的体积会变大,细胞质内出现的颗粒会明显增加。
- 产生这些形态学变化的主要原因,可能是因为在衰老的细胞内,蛋白质的降解体系受到抑制,从而导致蛋白质在衰老细胞的胞浆内持续地积累,以及衰老细胞中高尔基体和溶酶体增多造成的。
- 需要注意的是,上述这些典型性的细胞形态学变化并非绝对的。因为在不同类型的细胞内诱导衰老所形成的形态变化是不同的。而且即便是同一类型的细胞,由不同原因而导致的细胞衰老现象,表现出来的形态学变化也是不同的。
端粒缩短
- 端粒缩短现象不算是一个很好、很可靠的细胞衰老的标志物,只能说具有一定的参考价值。
- 端粒是位于生物体内染色体末端的一段特殊的DNA片段。在这段片段中含有许多简单的重复序列。在正常情况下哺乳动物体细胞中,可以合成端粒的端粒酶的活性是受到抑制的,因此随着细胞分裂次数增加,染色体末端的端粒长度不断地缩短,细胞分裂和DNA的复制能力越来越受到限制。
- 检测端粒的缩短现象是细胞衰老的特征之一。检测端粒长度的经典方法是Southern限制性内切酶酶切产生末端片段。但是这种方法所需样品量多,操作费时费力。
- 更加需要注意的是,有些过表达了 p16 的细胞中并没有发生衰老现象,但是有时也可以检测到端粒缩短和端粒的功能紊乱。
- 而且,由应激引起的细胞衰老现象中并没有端粒缩短现象的出现
衰老相关异染色质集落的形成SAHF
- 细胞要具备正常的生理功能和机能,依赖于细胞基因组的完整性。在某些外界刺激作用下,例如紫外线照射、化疗药物处理或者过氧化物刺激之后,细胞内产生多种信号,增加了细胞内活性氧自由基ROS的水平,或者引起了细胞内DNA损伤、DNA片段丢失、DNA链断裂、染色体移位等等,从而导致发生衰老的细胞内,产生衰老相关异染色质集落(senescence associated heterochromatic foci,SAHF)。
- SAHF现象在衰老的细胞内主要表现为细胞内染色质凝集,DNA凝结成散在的、致密的、大小不均一的异染色质颗粒。可以在细胞内通过DAPI染料对衰老细胞的DNA进行染色标记,在荧光显微镜下可以观察到在衰老细胞内有这种异染色质集落的形成现象。
- 有观点认为,SAHF的形成和p16信号通路密切相关。在SAHF的形成和维持过程中,组蛋白的甲基化修饰以及组蛋白相互结合蛋白的一系列变化。
- 在发生衰老的细胞内H3组蛋白第9位赖氨酸的三甲基化(H3K9me3)水平会提高;
- 异染色质蛋白1(heterochromatin protein 1,HP1)、高迁移率A族蛋白(high-mobility group A protein, HMGA)和组蛋白甲基转移酶 Suv39h1(suppressor of variegation 3-9 homolog 1)等分子的表达都有变化。
- 这些分子及H3K9的甲基化水平的变化,都可以作为检测细胞发生衰老的标志。
衰老相关分泌表型SASP
- 衰老的细胞除了表型出一定的特征性的细胞和分子的表征之外,还可以分泌一系列细胞因子,这些细胞因子通过自分泌或者旁分泌的方式,作用于衰老细胞本身或者作用于衰老细胞周围的其他细胞,从而改变了衰老细胞及其周围的微环境。
- 细胞由于发生衰老现象从而分泌的一系列的细胞因子,并改变衰老细胞自身或者周边微环境的现象,就被称为衰老相关分泌表型(senescence associated secretory phenotype,SASP ),或者被称为衰老信息分泌组(senescence-messaging secretome,SMS)。
- 衰老细胞可以分泌的细胞因子很多,例如,胰岛素样生长因子(insulin-like growth factor,IGF)、转化生长因子β(transforming growth factor-β,TGF-β)、白介素(interleukin,IL)、I型纤溶酶原激活物抑制剂(Plasminogen activatorinhibitor 1,PAI-1)等等。其中,IL-6(白介素-6)是非常重要的由衰老细胞所分泌的细胞因子。
- IL-6 的分泌直接由DNA 损伤信号通路调控,被认为和细胞衰老密切相关。
- 衰老细胞通过SASP机制从而将衰老细胞和其周围的细胞微环境联系在了一起。尤其在肿瘤发生过程中,SASP 机制被认为具有双重的调控作用。
- 一方面发生衰老的细胞通过SASP 机制,加速衰老细胞的生长停滞,促进机体启动免疫系统清除衰老的细胞,避免肿瘤的形成
- 另一方面,衰老细胞通过SASP机制所分泌的细胞因子通常可以引起炎症反应,参与破坏组织结构的完整性,甚至诱导细胞发生上皮间质转化EMT现象,也可能促进细胞癌变和癌细胞的增殖。
- SASP现象只能作为判断衰老的一个辅助标准,因为一些长期细胞或者病毒的感染、或者伤口愈合过程以及肿瘤发生过程中,受到感染或者发生癌变的细胞,同样可以分泌细胞因子,这类因子和衰老细胞通过SASP机制而分泌的细胞因子很相似。仅仅只是通过分析由细胞所分泌的细胞因子的种类,是不能准确判定细胞是否发生衰老现象的。
衰老相关分子的变化
- 在衰老过程中会有一些特异性的分子标志物的表达变化,通过检测这些特异性的分子标志物,可以对衰老现象进行鉴别。
- 需要特别说明的是,即便检测了衰老相关的分子标志物,也需要和其他衰老相关的特征相互结合,才能准确判定细胞衰老现象。任何依靠单一指标的检测,都是不全面的。
- 无论是采用经典的方法,还是最新的技术,细胞衰老的各种特征不是绝对特异的。不能仅仅根据细胞衰老的某一个特征来检测和判断细胞的衰老现象。因此,在研究细胞衰老现象时,需要根据细胞的多种特性综合而全面地分析,从而才能正确判断细胞是否发生了衰老现象。
衰老相关β-半乳糖苷酶活性的改变
- 这个分子的全称是senescence-associated β galactosidase,简写为SA-β-gal,它是目前应用最为广泛的细胞衰老的标志物之一,可以将发生衰老的细胞和处于分裂静息期或者是终末分化状态的细胞区分开来。
- 衰老的细胞内,SA-β-gal是由细胞溶酶体中的β-gal(β-半乳糖苷酶)衍生而来的。β-gal 在正常的人体细胞中主要表达在溶酶体中,最适pH范围是4-4.5。但是在发生衰老的细胞内,细胞的溶酶体开始膨胀,数量也增加,β-半乳糖苷酶在溶酶体中积聚,而且使得β-gal的最适pH范围发生了迁移,从正常细胞内的4-4.5迁移到衰老细胞内的pH6.0。因此,当细胞发生衰老时,β-gal可以在pH6.0时表现出高的酶活性。在检测衰老细胞时,可以人为给予β-gal的作用底物X-gal和pH6.0 的环境,结果衰老细胞可以在β-半乳糖苷酶的作用下,将底物转变为深蓝色的产物,使得发生衰老的细胞在普通光学显微镜下即可观察到。
- 检测SA-β-gal的方法操作简单,但是这种方法具有一定的局限性
- 首先,β-gal 在正常生理条件下也表达于一些非衰老的细胞中,例如破骨细胞、成熟的巨噬细胞等。
- 其次,正常情况下,在中性的pH环境中,β-gal 几乎不表达;但是在细胞接受强的电离辐射之后,在1 个小时内细胞内的β-gal 就可以表现出极高的酶活性,因此会给细胞衰老的判断带来极大的干扰。
- 通过检测SA-β-gal的方法鉴定细胞衰老,需要谨慎,尤其是研究溶酶体活动旺盛的细胞、或者研究DNA受到损伤的细胞、与炎症和肿瘤相关的巨噬细胞时,要防止误判。
和细胞周期相关的分子标志物
- 细胞衰老的主要的特征之一就是细胞永久性地退出细胞周期。因此很多细胞周期相关蛋白都在细胞衰老过程中,起到重要的作用,可以作为细胞衰老的标志物。
p16/p14
- 复习二十四型 Lesson 2,其中我们提到了一个基因CDKN2A,cyclin-dependent kinase inhibitor 2A,细胞周期素依赖性蛋白激酶抑制因子2A。CDKN2A基因编码了两个重要的细胞周期调控蛋白p14和p16。p14和p16是两个不同的蛋白,发挥不同的功能。p14和p16相比,有共同的外显子2和3,但是它们的外显子1是不同的。P14是外显子1β;p16是外显子1α。由于p14和p16的外显子1不同,而且发生了读码框的移位,导致p14和p16的蛋白序列也是不同的。
- p16的分子量是16kD,由156个氨基酸残基组成,大约75%的癌细胞株中都存在p16蛋白的缺失或者突变。在衰老的细胞内,p16调控了CDK4和CDK6,进而影响了Rb信号通路,阻断细胞周期,促使细胞发生细胞衰老现象,因而p16分子可以作为细胞衰老的标志物之一。
- 有研究发现,在通常情况下,p16蛋白在健康且没有应激压力反应的组织细胞内表达量很低,但是在衰老的细胞、受到损伤的细胞和初期的肿瘤细胞内,p16的表达会明显地升高,因此p16的表达具有明显的动态特性。
- 如果作为细胞衰老的标志物,p16具有较大的局限性。
- 第一,某些没有衰老的细胞,例如部分免疫细胞或者肿瘤细胞内,同样高表达p16。譬如,多次复制的T细胞内就积累了大量的 p16 蛋白,但是细胞却并不表达其他的衰老标志物,而且,分离出这类高表达p16的细胞后,如果给予这些细胞以适宜的细胞培养条件,细胞能重新进入细胞周期。由此可知,单纯检测p16蛋白的高表达不能准确判定细胞的衰老状态
- 其次,某些类型的衰老细胞中不表达或者低表达p16蛋白。所以,使用p16作为衰老标志物需要谨慎。
- CDKN2A基因所编码的另一个蛋白p14,可以作为p53 信号的上游信号通路,通过p14-MDM2-p53 信号通路,从而参与调控细胞的衰老过程。因此,有些学者认为,p14分子也可以作为细胞衰老的标志物之一。但是,老谈个人不推荐使用p14作为细胞衰老的标志物,因为p14作为调控p53信号通路的关键调控因子之一,广泛参与p53通路的调控。而p53通路的作用范围又是如此地广泛,会导致几乎所有发生p53通路发生变化的细胞内,几乎都可以检测到p14蛋白的变化。因此,检测p14的蛋白变化,根本不能表征细胞衰老的现象,只能带来干扰。
p16/p14
- 编码p21的基因是CDKN1A,cyclin-dependent kinase inhibitor 1A,细胞周期素依赖性蛋白激酶抑制因子 1A,其基因位于人6号染色体上,分子量21kD,由三个外显子编码的164个氨基酸残基组成。p21的表达受到p53的调控,并且参与了细胞周期和衰老相关的信号传递通路。
- 有研究发现,在小鼠中敲除CDKN1A基因后,小鼠细胞内出现G1-S和G2-M检查点的功能紊乱,从而导致小鼠体内产生更多的肿瘤。
- 进一步的研究发现,p16也同样介导了细胞的衰老现象。
细胞衰老相关信号通路
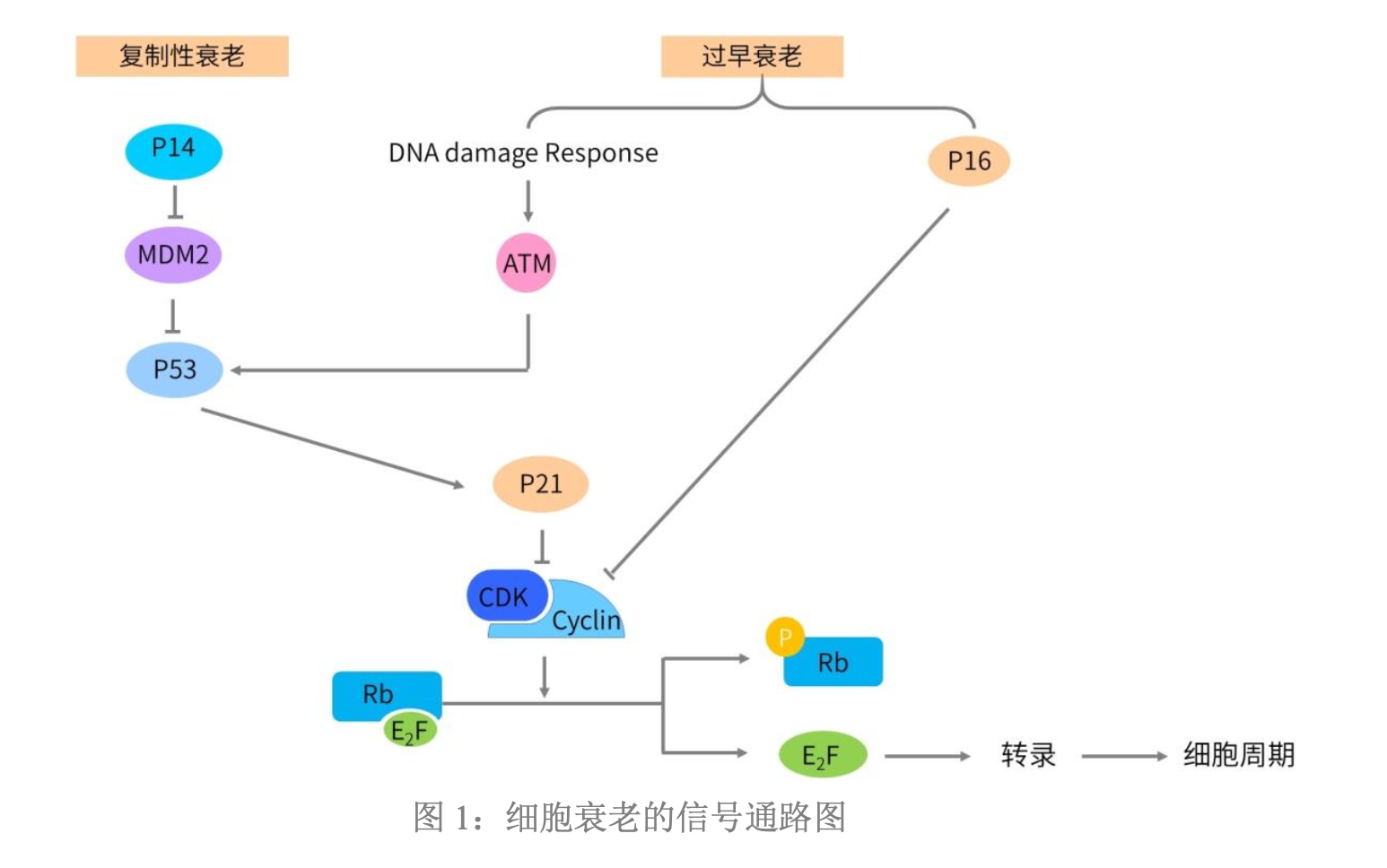
MDM2-p53相互作用
- 在正常的细胞内:MDM2和p53相互结合,并且抑制p53的功能和活性,促进p53的蛋白降解,从而始终在正常细胞内保持p53的低表达。
Rb通路
- 当细胞处于非分裂状态时,细胞内的Rb处于非磷酸化的状态,而且非磷酸化的Rb和转录因子E2F相互结合,抑制E2F转录的活性。
- 在细胞发生正常分裂时,生长因子或者有丝分裂原和相应的受体结合,通过信号通路传递促进cyclin基因表达;随后cyclin和相应的CDK相互结合,形成cyclin-CDK复合物,促进Rb蛋白的磷酸化,形成pRb。pRb与E2F分离,自由的E2F进入细胞核内,调控了下游基因的转录,这些受到E2F转录调控的基因产物促使细胞进入细胞周期。
复制性衰老的通路
- 首先,当细胞增殖和分裂的次数越来越多,细胞内染色体上的端粒末端的序列也不断地丢失,导致端粒的序列越来越短。当端粒缩短到无法维持染色体结构的完整性的时候,细胞内的p14被激活。
- 激活的p14能抑制MDM2的活性,从而间接地促进了p53的稳定并激活了p53 的活性。激活后的p53可以入核,调控下游基因的转录表达。
- 在p53的下游基因中,最重要的就是CDKN1A以及CDKN1A基因编码的p21蛋白。p21表达升高后,抑制了cyclinE/CDK2 复合物的活性,使得磷酸化的pRb蛋白转变为非磷酸化的Rb蛋白。非磷酸化的Rb蛋白重新结合转录因子E2F,使得E2F不能激活细胞周期所必需的基因表达,促使细胞发生G0/G1期细胞周期阻滞,无法进入S期,也无法启动染色体复制,抑制了细胞的分裂和增殖,最终启动细胞的复制性衰老。
- 当细胞发生衰老现象时,通过p14-MDM2-p53-p21-pRb通路,促使细胞退出细胞周期,并启动细胞衰老的进程,在整个过程中,p14的激活是整个通路的上游起始诱因,通过下游的p53通路和Rb 通路,最终实现细胞退出细胞周期并开始衰老的整个过程
过早衰老的通路
- 当细胞受到外界刺激而发生过早衰老时,下游的信号通路和复制性衰老的通路略有差异。
- 在细胞受到外界刺激时,往往会导致细胞内的DNA发生损伤,从而引起DNA 损伤反应(DNA damage response),在这个过程中ATM基因被DNA损伤反应所激活。
- ATM 激活后,促进p53的磷酸化,并激活p53的功能。随后p53调控下游p21的转录,使得p21表达升高。表达升高后的p21通过Rb通路,抑制 E2F,促使细胞退出细胞周期并开始衰老过程。
- 在这条通路中,除了p53由ATM激活之外,p53及其下游通路和复制性衰老过程中的通路是类似的。差别只是在激活p53的起始诱因上的差异:当细胞发生复制性衰老时,细胞内p14激活,然后通过MDM2间接地激活p53;而当细胞发生过早衰老时,细胞内发生DNA损伤反应,先激活ATM再激活p53通路。
- 在细胞发生过早衰老时,除了上面提到的ATM-p53-p21-Rb 通路之外,发生过早衰老的细胞还能促进细胞内p16的表达上升和功能激活。p16是一种CKI,也就是CDK4和CDK6的活性抑制剂。当p16表达升高后,通过抑制CDK,进而抑制了CDK/cyclin复合物,从而促使磷酸化的pRb转变为非磷酸化的Rb。非磷酸化的Rb结合转录因子E2F,抑制了E2F。
细胞衰老现象与肿瘤
- 很早之前,就有研究人员认为细胞衰老是防止细胞发生癌变的第三道保障。第一道保障是细胞内DNA 的损伤修复机制;第二道保障是诱导细胞凋亡的能力;第三道保障是诱导细胞发生衰老。长久以来,大多数的研究人员都认为细胞衰老对于肿瘤发生具有抑制作用。
细胞衰老对于肿瘤发生的抑制作用
- 细胞衰老过程中,Rb和p53蛋白在诱导细胞发生衰老时起到了至关重要的作用。而Rb蛋白和p53蛋白又是非常关键的抑癌基因,抑制了细胞发生癌变的过程。很多肿瘤的发生都是由于抑癌基因的失活而导致的。
- 研究表明,Rb和p53蛋白的活性增强,可以启动细胞衰老过程。因此,如果细胞要发生癌变,那么规避衰老现象是细胞发生癌变的必要条件。现在越来越多的研究,开始关注如何启动细胞衰老现象从而抑制肿瘤的发生发展。
- 有研究发现,在PTEN基因失活的小鼠前列腺模型中,可以在癌前病变的小鼠组织中检测到细胞衰老现象,但是在发生了恶性前列腺癌的小鼠组织中却没有检测到细胞衰老现象。而这种差异的关键在于p53信号是否激活。
- 另有研究发现,在小鼠模型中高表达K-Ras^v12 癌基因,可以诱导癌前病变、多发腺瘤,甚至是恶性的腺癌。在癌前病变和腺瘤组织中可以检测到细胞衰老,以及p16、Dec1以及HP-1等衰老标志分子,但是在腺癌细胞中不能检测到细胞的衰老现象。这些研究结果都提示激活的癌基因在小鼠体内诱导细胞衰老,但是由于少数细胞内的突变积累导致极少数的细胞突破了衰老防线,并进一步发展为恶性肿瘤。
细胞衰老对于肿瘤发生的促进作用
- 体内的衰老现象是一种重要的抗肿瘤防卫机制,但是由于衰老细胞仍然保留基本的细胞代谢过程,同时发生衰老的细胞可以产生各种活性蛋白和细胞因子(也就是SASP现象)也可能对肿瘤发生产生了促进的作用。
- 衰老细胞可以不断积累产生各种细胞炎性因子以及上皮生长因子,从而对衰老细胞周围的微环境的结构和功能造成了破坏,因此衰老细胞可以通过SASP现象而间接地对细胞造成损伤,甚至诱导细胞恶性增殖并发生癌变。
- 衰老不足可能引起肿瘤的发生,但是过度地衰老也可以导致肿瘤的发生。如何平衡这两者的矛盾,需要我们不断深入进行研究。
- 目前学术界提出了“拮抗性多效假说”(antagonistic pleiotropy hypothesis),认为衰老在早期可以保护机体不至于发生肿瘤,而到了机体的老年时期则促进衰老表型,促使肿瘤的形成。当然这种理论,目前还是一种假说,需要更深入的研究和确实的实验证据。


